

Versión 1.
CONCEPTOS BÁSICOS DE GENÉTICA CUANTITATIVA
Los genes que determinan los rasgos fenotípicos cuantitativos (los que presentan una variación continua en la población: estatura, peso, inteligencia … ) tienen la misma entidad y se trasmiten de padres a hijos según los mismos principios de trasmisión de la herencia descubiertos por Mendel.
Rasgos cualitativos/fenotipos discretos. Son cualitativamente diferentes entre sí (en el caso del color de los guisantes, se comprueba que son o amarillos o verdes; en el caso del factor rh, o se es positivo (rh+ = presencia de antígeno) o negativo (rh– = ausencia de antígeno), sin valores intermedios.
Rasgos cuantitativos/fenotipos continuos. Son cuantitativamente diferentes, sólo se diferencian entre sí por el valor cuantitativo que muestra el rasgo en cada individuo (un individuo puede ser 1 centímetro más alto o más bajo que otro, o pesar una décima de gramo más o menos que otro)
En términos genéticos, la explicación de la diferencia entre rasgos cualitativos y cuantitativos se halla en el hecho de que los rasgos cuantitativos están determinados (normalmente) por varios genes, cada uno con dos o más alelos, donde cada alelo contribuye con una cierta cantidad al fenotipo observado. Mientras que los rasgos cualitativos son monogénicos. Puede darse el caso de que un rasgo monogénico sea cuantitativo, es lo que en genética mendeliana se llama dominancia o herencia intermedia.
Los experimentos que realizo Mendel con plantas de guisantes nos llevan a la conclusión obvia y evidente es que la altura de las plantas de guisante es un rasgo discreto, cualitativo, mendeliano o monogénico.Algo similar pero con plantas de tabaco lo llevó a cabo un tal Josef Gottlieb Kolreuter (1733-1806) hacia 1760, sólo que sus resultados fueron por completo diferentes, al menos en apariencia.
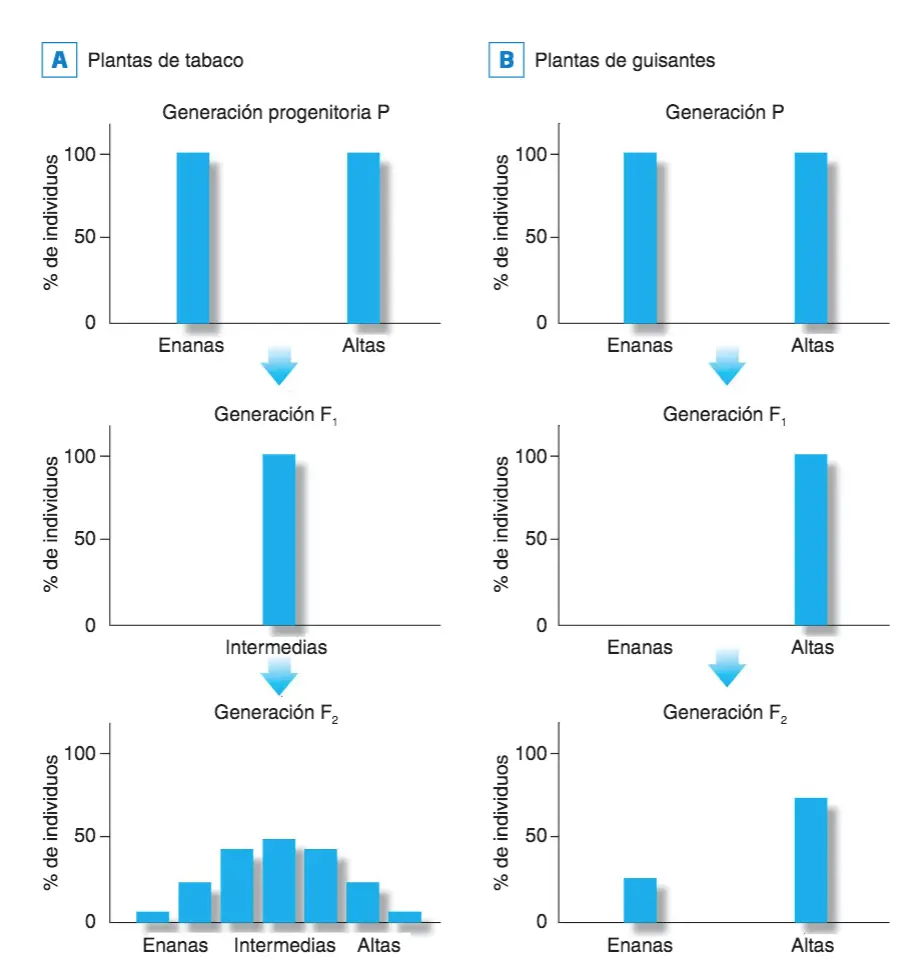
A. Cruzamientos mendelianos de plantas de tabaco donde la generación P ha sido seleccionada por la altura utilizándose como progenitores individuos con valores extremos en el rasgo fenotípico altura; el hecho de que todos los miembros de la generación F, sean de una altura intermedia y los de la generación F2 se distribuyan según la curva normal en cuanto a este rasgo fenotípico está indicando que la altura en esta planta de tabaco es un rasgo cuantitativo poligénico. B. Como ya sabíamos por los experimentos de Mendel, este mismo rasgo, la altura, en los guisantes es un rasgo monogénico, puesto que se cumple en él la segunda ley de Mendel.
Herman Nilsson-Ehle (1873-1949) en 1908 realizo un experimento: cruzó trigo de grano rojo con trigo de grano blanco. La diferencia entre los resultados de Mendel y los de Nilsson-Ehle se halla en los fenotipos
de los restantes granos de trigo, porque, el color va del rosa claro al rojo pasando por valores de color intermedios en unas proporciones muy características: 1/16 de rojo intenso, 4/16 de rojo desvaído, 6/16 de rosa y 4/16 de rosa pálido; sólo falta añadir el 1/16 de granos blancos para completar el total de 16/16.

¿Cómo explicar estas frecuencias fenotípicas, tan diferentes en apariencia de las mendelianas? Hacemos una Tabla de Punnett. Para describir el concepto de cantidad damos el valor de 1 a los alelos que aportan color (A = 1 y B =1 ) y el valor 0 a los que no aportan color (a= 0 y b =0); en cada casilla se representa el genotipo y la cantidad de color del fenotipo representada por la suma de los valores que se
les ha dado a cada alelo. En genética, a los alelos cuyo valor se suma al de otros para explicar el fenotipo reciben el nombre de alelos aditivos.
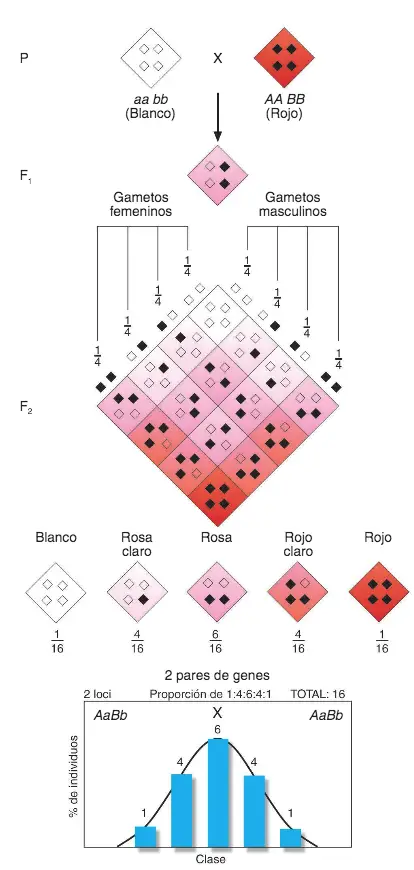
Resultados de los cruzamientos mendelianos de plantas de trigo a partir de una generación P donde los progenitores son razas puras con va lores extremos del rasgo color de la semilla (roja/blanca).
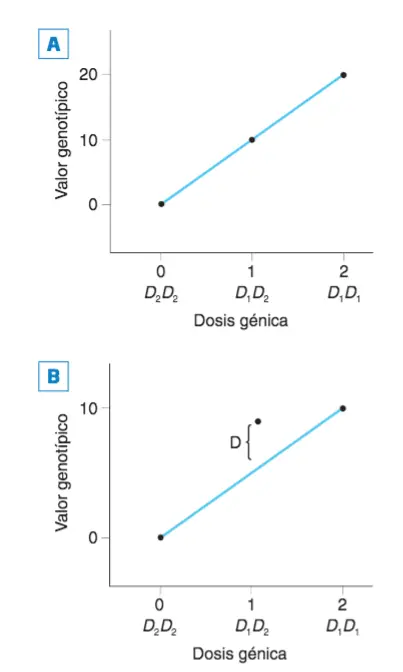
Valor genético aditivo y efecto de dominancia. (A). Suponiendo que el gen D es un gen con dos alelos, D1 y D2, de los que el primero, D1, posee un valor aditivo de 10 mientras que el otro, el D2, tiene un valor aditivo de 0, podemos representar, como se hace en la figura, cuál será el efecto sobre el fenotipo (valor genotípico) de un determinado genotipo en función de la dosis alélica (o número de copias de un alelo que porta un genotipo). En el eje de abscisas (horizontal) se representa la dosis génica del alelo D1: puesto que el valor aditivo de este alelo es de 10, cuando su dosis génica es de 2, el valor genotípico será de 20; puesto que el valor aditivo del otro alelo es de cero, el valor genotípico del heterocigoto es de 10 (dosis génica de D2 igual a 1; dosis génica de 0, igual a 1; ver texto). (B). Cuando la dominancia de un alelo sobre otro es completa, los valores genotípicos no se ajustan a lo esperado de acuerdo con la dosis génica: puesto que basta una copia del alelo dominante para obtener el efecto máximo sobre el fenotipo, el valor genotípico será el que se representa en la gráfica. A esto es a lo que se llama efecto de dominancia (podría ocurrir también que el alelo dominante fuera el que no aporta valor al fenotipo, en este caso el alelo D2, en cuyo caso, el genotipo heterocigoto,D1D2 presentaría un valor fenotípico de 0, igual que el genotipo D2D2) .
Conceptos clave:
Valor aditivo. Es el valor numérico de los alelos en función de la cantidad que cada uno aporta al fenotipo final.
Dosis génica o número de veces que aparece un alelo de un gen en un genotipo. El concepto de dosis génica (gene dosage) hace referencia al número de copias de un gen: se habla de dosis de la misma manera que se habla de dosis cuando se trata de administrar una medicina. Obviamente, la dosis génica en un individuo dado podrá ser de 2, cuando es homocigótico para ese alelo, o de 1 si es heterocigótico.
Valor genotípico. Es el resultado de sumar la dosis génica de cada alelo multiplicada por el valor aditivo
de cada alelo. P. ej., en el caso del color de los granos de trigo. La dosis génica del alelo A de las plantas AABB = 2: dosis de A = 2; multiplicando el valor numérico de la dosis (2) por el valor aditivo del alelo A (1) obtenemos el valor genotípico, en este caso 2 (2A = 2 X 1).
Según los planteamientos de la genética cuantitativa, Davenport (1913) planteó el mismo modelo del
color del grano de trigo para explicar las variantes de color de la piel humana en Jamaica.
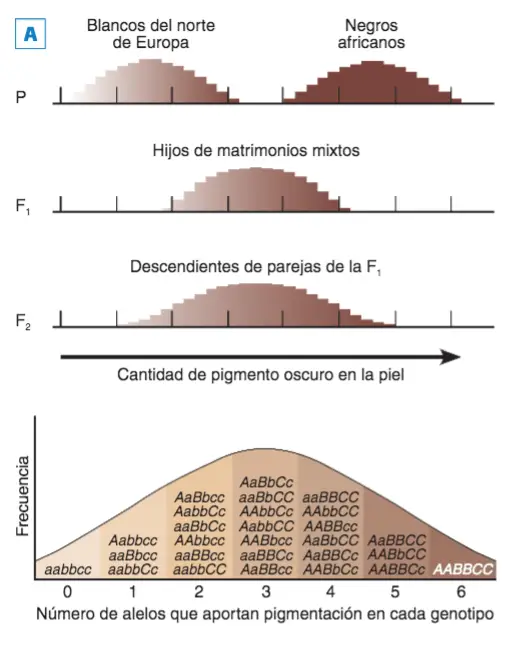
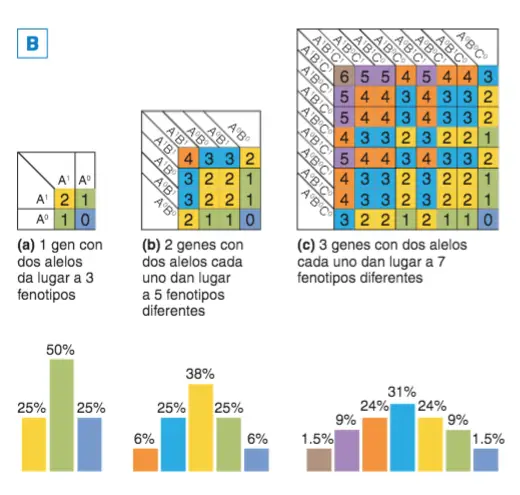
A. Distribución de la pigmentación de la piel en grupos humanos. Hipotética distribución de la pigmentación de la piel asumiendo tres genes con dos alelos donde sólo los designados con mayúscula aporta pigmentación; en este caso, encontramos 7 fenotipos diferentes. B. Representación de los genotipos y fenotipos posibles partiendo de 1, 2 o 3 genes con dos alelos aditivos cada uno.
Identificación de los genes aditivos: locus de rasgo cuantitativo
A diferencia de lo que ocurre con los rasgos monogénicos (han podido ser ubicados en cromosomas concretos), la localización de los genes aditivos asociados a rasgos cuantitativos es una incógnita, igual que lo referente a rasgos conductuales, normales o patológicos. No obstante, gracias al proyecto genoma humano que ha permitido cartografiar todo nuestro genoma, y probable que algún día se identifique el loci donde se alojan estos genes aditivos, los llamados loci de rasgo cuantitativo.
Estas investigaciones reciben el nombre de Estudios de Asociación Genómica Amplia (Genome-Wide Association Studies (GWAS), y requieren un número muy elevado de sujetos para encontrar loci que presenten una asociación o correlación con el rasgo de interés.
GENÉTICA CUANTITATIVA: HEREDABILIDAD
La variabilidad genética es un factor a tener muy en cuenta se quiere explicar la conducta. El ambiente es otro gran factor a tener en cuenta. La conducta en cuanto fenotipo es, evidentemente, el resultado de las influencias conjuntas de los genes, y el ambiente.
¿Se puede averiguar en qué medida un rasgo psicológico, o las adicciones, está determinado por los genes y en qué otra medida lo está por el ambiente? Con lo que sabemos sobre el mecanismo de regulación de la expresión génica, cualquier rasgo está determinado por los genes y por el ambiente. Sin embargo, si mantenemos constante el ambiente, es obvio que las diferencias que encontremos en la población (variabilidad) sólo cabrá atribuirlas a diferencias genéticas, de la misma forma, si sabemos que en una población dada el genotipo es el mismo para todos los individuos, las diferencias que encontremos, si las hubiere, sólo se podrán atribuir al ambiente.
¿Cuánto de la variabilidad observada en un rasgo fenotípico cualquiera puede atribuirse a los genes y cuánto al ambiente? La proporción de variabilidad atribuible a diferencias genéticas recibe el nombre técnico de heredabilidad y es el concepto más importante de la genética cuantitativa. La heredabilidad es un valor numérico entre 0 y 1; ese valor numérico representa la proporción de la variabilidad del rasgo fenotípico atribuible a los genes; en contraposición, la ambientalidad es la proporción de la variabilidad atribuible a factores ambientales. Llamaremos H2 a la heredabilidad y A a la ambientalidad.
El parámetro que cuantifica la variabilidad de un rasgo en una muestra es la varianza. La varianza del rasgo tal como se presenta recibe el nombre de varianza total o varianza fenotípica (VT) . La VT es igual a la suma de la varianza genética (VG) más la varianza ambiental (VA), más la varianza resultante de la interacción entre genotipo y ambiente (VGA). En los estudios de genética cuantitativa humana esta interacción se suele obviar porque no es posible calcularla eficazmente.
VT = VG + VA
- VG = fracción de la varianza fenotípica (varianza total) debida a diferencias genéticas entre los individuos de la población.
- VA= fracción de la varianza fenotípica debida a diferencias en las condiciones ambientales a las que se han visto expuestos los individuos de la población.
A partir de la formula de la varianza fenotípica (VT) obtenemos la fórmula para calcular la heredabilidad:
H2 = VG / VT = H2 = VG / (VG + VA)
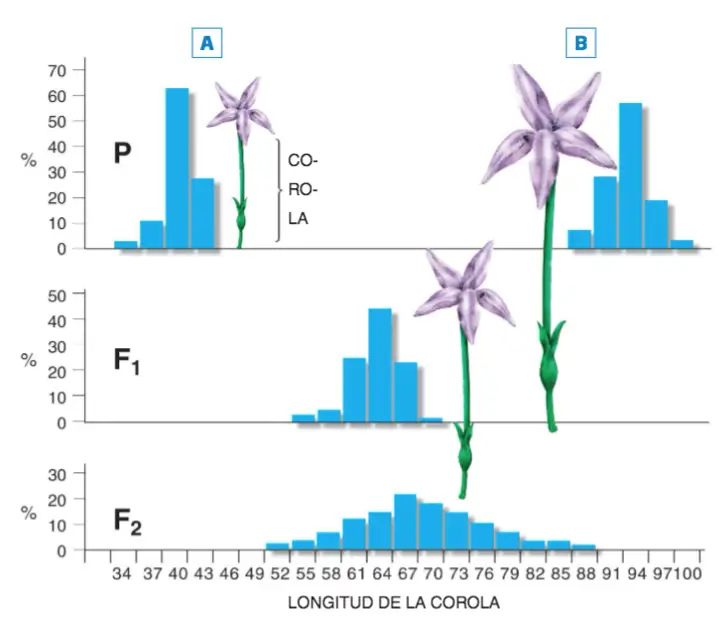
La longitud de la corola de la flor de la planta de tabaco Nicotiana longiflora es un rasgo cuantitativo, tal como lo demuestra el hecho de que la longitud de las corolas de las flores de la generación F2 se distribuye según la curva normal y no según las proporciones mendelianas características de los rasgos cualitativos.
Investigación realizada por E.M. East con plantas de tabaco. La longitud de la corola de una variedad de la planta de tabaco Nicotiana longiflora era ostensiblemente más corta (3 7-43 mm: variedad A) que los de otra variedad de la misma planta (9 1- 97 mm: variedad B). La varianza observada en cada una de estas dos variedades fue de 8.76 (VA = 8.76), la misma varianza que se observó en la generación F1 resultante del cruce entre las dos variedades primitivas (P). Puesto que las dos variedades parentales A y B son homocigóticas (razas puras) y todos los miembros de la generación F1 son genéticamente idénticos, la varianza de 8.76 sólo puede deberse a la influencia ambiental (VA). Cuando se cruzaron entre sí los miembros de la generación F1 para obtener la generación F2, se comprobó que la varianza total (VT) era de 40. 96 (VT = 40. 96). Conociendo la VA (8.76) y la VT (40.96), podemos calcular la varianza genética (VG).
VT = VG + VA; 40.96 =VG+ 8.76; VG= VT– VA; 40.96 – 8.76 = 32.20
Si queremos calcular el valor de la heredabilidad (H2), bastará con que apliquemos la fórmula: H2 = VG / VT
H2 = 32.20 / 40.96 = 0.79
0.79 no significa que el 79 % de la longitud de las flores de Nicotiana longiflora se deba a los genes y el 21 % restante al ambiente; lo que la heredabilidad representa es la proporción o porcentaje de la variación fenotípica observada entre los individuos de la población que se debe a las diferencias genéticas entre esos mismos individuos.
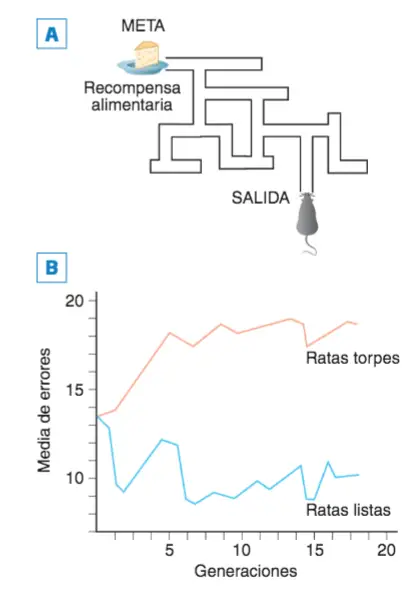
En 1924 Tolman (1886-1959) empezó a criar selectivamente ratas en función de su eficacia para aprender un laberinto usando comida como refuerzo: sus resultados, completados por Tryon, demostraron que la capacidad de aprendizaje, en las ratas, se ve notablemente influido por factores genéticos, puesto que bastaron ocho generaciones para conseguir que todas las ratas «listas» fueran más rápidas en aprender la tarea que la más lista de las ratas «torpes».
A. Plano del laberinto que tenían que aprender las ratas de Tolman y Tyron para alcanzar el refuerzo (la comida). B. Gráfica de resultados obtenidos por generaciones de ratas seleccionadas en función de la facilidad o dificultad con que aprendían el laberinto (el aprendizaje se puede medir por el número de veces que necesita una rata para alcanzar la meta en el menor tiempo posible y también, como es el caso, por el número de errores en tomar el camino correcto, el más directo).
Una manera directa de calcular la heredabilidad consiste en evaluar el efecto de la cría selectiva entre dos generaciones. Experimento de selección realizado por Johannsen (1857-1927) en 1903 con alubias comestibles del género Phaseolus.
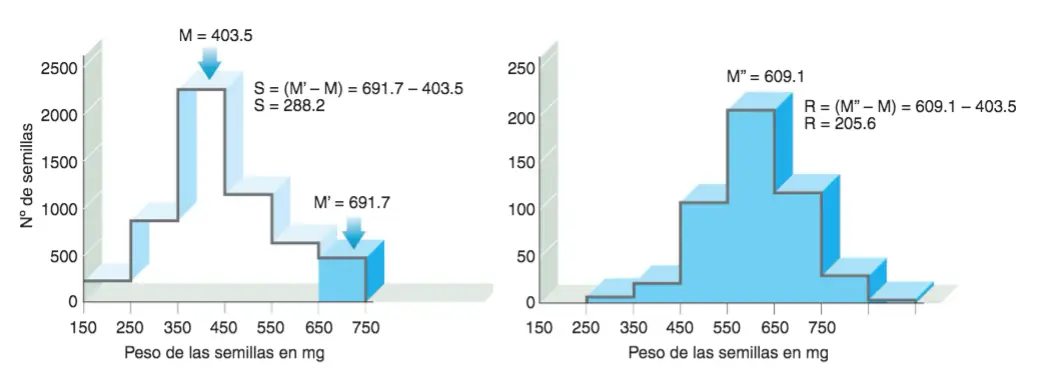
Demostración de la heredabilidad a partir de estudios de cría selectiva. M: media de la población original; M’: media de la subpoblación de individuos seleccionados como reproductores; M»: media de la población descendiente del grupo de individuos seleccionados como reproductores; S: diferencial de selección; R: respuesta a la selección.
En una población de semillas cuya media era de 403.5 mg, se seleccionaron para la reproducción semillas con una media de 691.7 mg. A la diferencia entre la media de la población general (M) y la de las semillas elegidas (M’) se la designa con el nombre de diferencial de selección y con la sigla S: S = (M’ – M) = 691.7-403.5 = 288.2. La media del peso de las alubias descendientes M» resultó ser de 609.1; con lo que la llamada respuesta de selección o Res de 205.6 (R = (M» – M) = 609.1 – 403 .5 = 205.6). Para calcular la heredabilidad, basta con dividir R por S:
H2 = R / S = (M»- M)/(M’ – M) = 205.6 / 288.2 = 0.713
Llegados a este punto, es conveniente volver sobre la fórmula general de la heredabilidad: H2 = VG / VT
Hasta ahora hemos estado asumiendo que la heredabilidad depende de la varianza genética aditiva (VGa), es decir, hemos asumido que la parte de la varianza total explicada por factores genéticos sólo era debida al hecho de que sólo existen dos variantes del gen de que se trate (genes aditivos dialélicos).
Por lo que sabemos de las leyes de Mendel, esto no tiene por qué ser así. Cuando se da interacción entre genes que ocupan diferentes loci, los resultados de los cálculos no se corresponderán estrictamente con los esperados a partir de una varianza meramente aditiva.: es el caso de la epistasia (epistasis) A la varianza debida al efecto de dominancia la designaremos como VGd y a la varianza atribuible a la epistasis la llamaremos VGi (i por interloci).
Entonces, la ecuación para calcular la heredabilidad, H2 = VG / VT se transforma en:
H2 = (VGa + VGd + VGi) / ((VGa + VGd + VGi + VA)
Esta fórmula corresponde a la heredabilidad en sentido amplio (H2). La heredabilidad atribuible específicamente a la varianza genética aditiva, la que explica el éxito de la cría selectiva, recibe el nombre de heredabilidad en sentido estricto (h2) y su fórmula es
la siguiente:
h2 = VGa / VT
Heredabilidad en sentido estricto (h2). «La heredabilidad en sentido estricto se asume que la variabilidad observada es debida sólo a los efectos aditivos». aLF ED
Heredabilidad en sentido amplio (H2). «La heredabilidad en sentido amplio refleja la variabilidad debida a todos los tipos de acción de los genes; efectos aditivos, efectos de dominancia, efectos de epistasia y efectos de interacción de genes por ambiente». aLF ED
Esta distinción es sumamente importante para la genética de la conducta humana, donde la varianza genética sólo se puede evaluar a partir del parentesco genético, dado que en cada parentesco se combinan de forma diferente los tres componentes de la varianza genética, el aditivo, el debido a la dominancia y el atribuible a la epistasia.
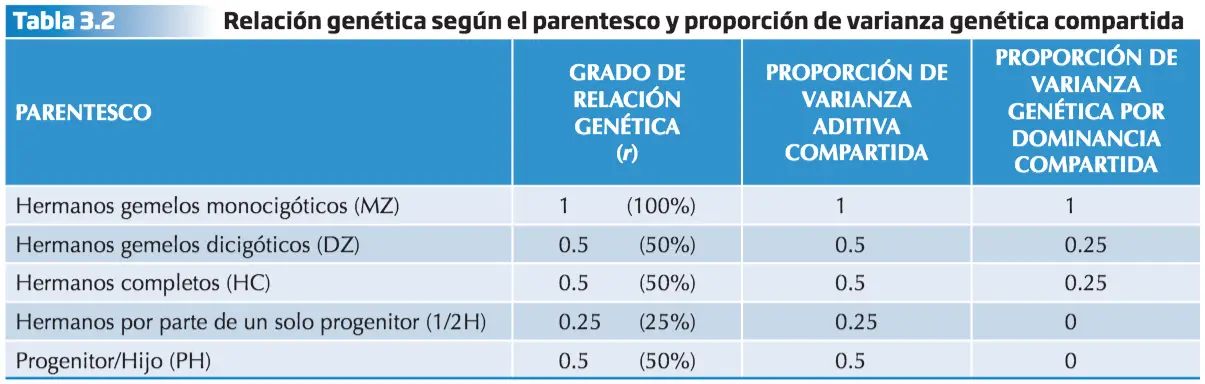
La epistasia sólo es compartida al 100% entre hermanos monocigóticos. En el resto de los casos se considera mínima. Padres e hijos no pueden compartir la relación de Dominancia, por ello sólo es tenida en cuenta entre hermanos.
En la medida en que asumamos que los efectos del ambiente se distribuyen al azar con respecto a los valores genotípicos y que no se da interacción genotipo/ambiente, estos valores de la covarianza sirven para calcular la heredabilidad. Asumir que el efecto del ambiente se distribuye al azar en el caso humano no deja de ser una fantasía.
En los estudios con humanos, la heredabilidad se calcula a partir del coeficiente de correlación (o de regresión) Se parte del supuesto, que no siempre puede cumplirse, de que la influencia del ambiente ni correlaciona con la de los genes, ni interactúa con ellos. Si esto fuera así, la heredabilidad tendría el valor de la correlación entre gemelos monocigóticos o idénticos: en este caso, estaríamos hablando de heredabilidad en sentido amplio puesto que comparten el 100% de los alelos, tanto los aditivos como los no aditivos
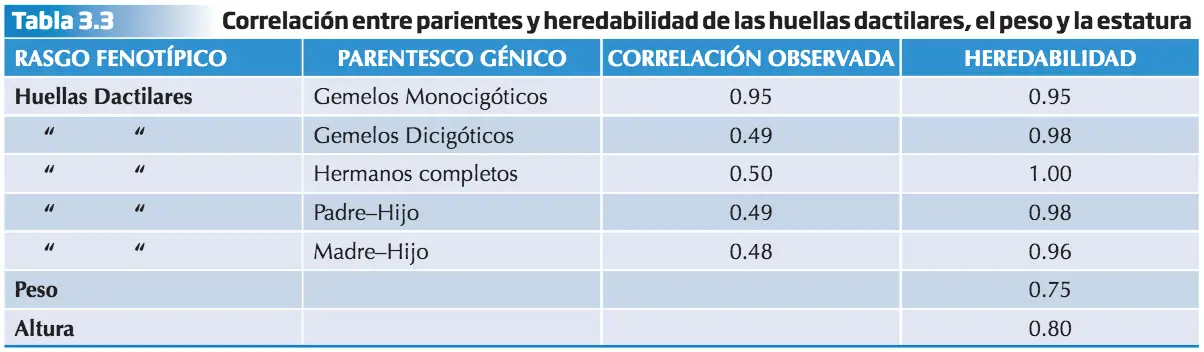
El valor de la Heredabilidad calculado a partir del parentesco es el valor de la correlación entre los parientes considerados dividido por el grado de parentesco genético. Tomemos como ejemplo los datos obtenidos en las tablas 3.2 y 3.3. Para calcular la Heredabilidad del rasgo “huellas dactilares” en gemelos monocigóticos:
H2 = Valor de correlación / Grado de parentesco genético = 0.95 / 1 = 0,95
Cuando calculamos la heredabilidad a partir de la correlación:
- Entre hermanos (completos, gemelos homocigóticos o dicigóticos), la heredabilidad obtenida es en sentido amplio (H2).
- Entre padre/madre e hijo, la heredabilidad obtenida es en sentido estricto (h2), ya que los efectos de dominancia y Epistasia no influyen.
Si el caso de la dermatoglifia, el ambiente no nos parece relevante, no creo que podamos decir lo mismo cuando nos refiramos a rasgos psicológicos o conductuales del tipo de la inteligencia, la memoria, la agresividad, la esquizofrenia, la depresión, la drogadicción, etc.
¿Qué hacer entonces para corregir los hipotéticos errores debidos a la covarianza entre genes y ambiente y a la interacción entre ellos? Falconer (Falconer y Mackay, 1996) ha desarrollado una serie de fórmulas que permiten calcular, en teoría, tanto la heredabilidad como la ambientalidad basándose en dos factores que se sabe a priori que están influyendo en el fenotipo:
- Según el parentesco:
- gemelos monocigóticos.
- gemelos dicigóticos.
- Según la crianza:
- Criados juntos
- Criados por separado.

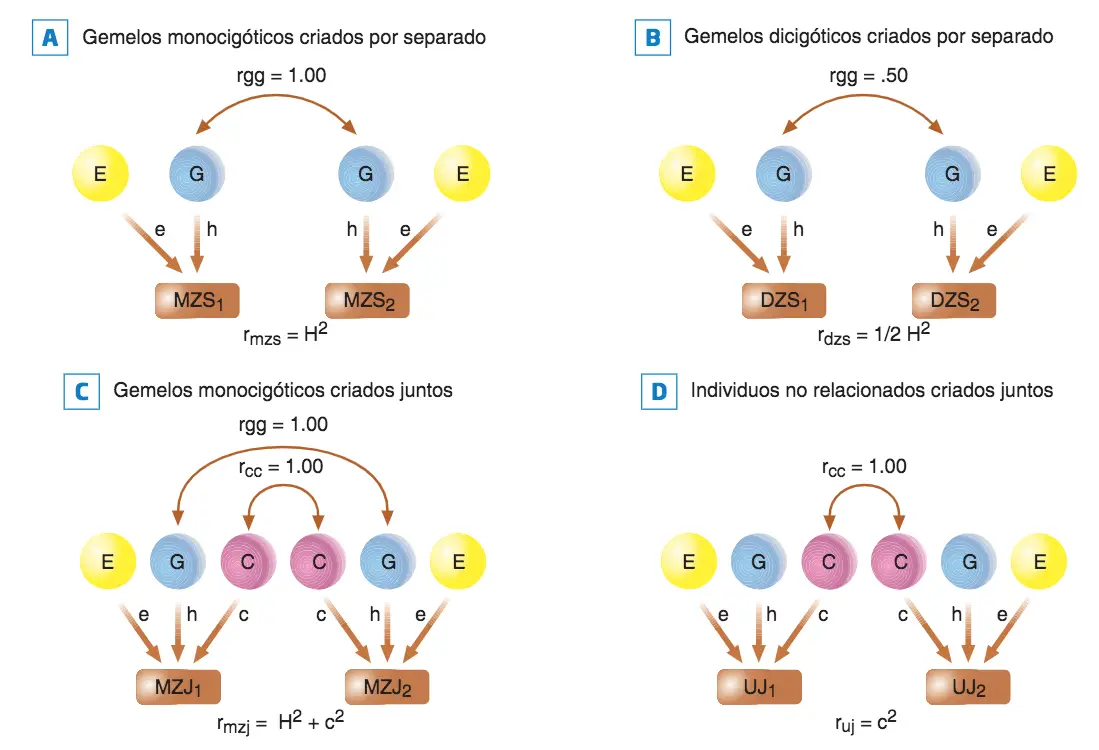
Diagramas de representación del valor teórico de los factores que intervienen en la correlación entre individuos: (A) Gemelos monocigóticos criados por separado (MZS), (B) gemelos dicigóticos criados por separado (DZS); (C) Gemelos monocigóticos criados juntos (MZJ); (D) Individuos no relacionados criados juntos (UJ). g: genético; e: ambiente no compartido; h: herencia; e: ambiente compartido; u: no relacionado genéticamente; rgg: grado de parentesco genético; rcc: grado en el que se comparte el ambiente; rmzs: correlación entre MZS; rdzs: correlación entre DZS; rmzj: correlación entre MZJ; ruj: correlación entre sujetos que crecen juntos sin ser parientes genéticos; H2 : heredabilidad en sentido amplio; c2 : ambiente compartido.
Cuando no se puede contar con gemelos criados por separado, Falconer nos ofrece otra estrategia para obtener la heredabilidad:
H2 = 2 (rMzj – rDZj)
Una segunda fórmula propuesta por Falconer sirve para obtener el posible efecto del ambiente compartido c2:
c2 = (2 · rDZj) – rMZj
Falconer también ofrece una fórmula para calcular la ambientalidad específica (no compartida (e)):
e = 1 – rMZ
La principal limitación del significado de la heredabilidad consiste en que su aplicación se limita al momento y a la población en la que se ha obtenido; es decir, está sujeta a la ambientalidad peculiar perteneciente a ese momento y a esa población. Para comprender cómo puede ser eso así, nos serviremos de los estudios de Cooper y Zubek (1958) sobre la genética del aprendizaje de laberintos en las ratas seleccionadas por Tolman y Tryon.
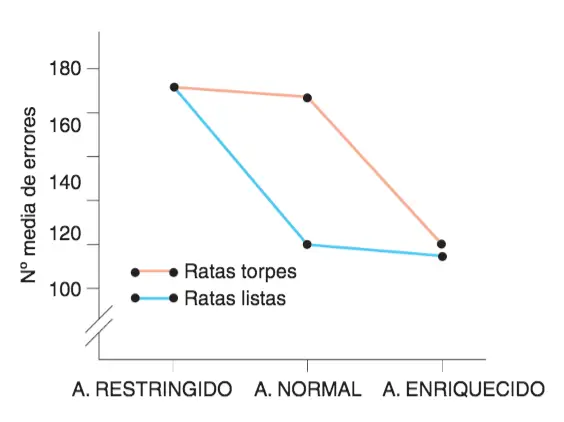
La heredabilidad de la capacidad para aprender laberintos sólo se puso de manifiesto en condiciones normales desapareciendo cuando el ambiente durante el desarrollo fue enriquecido o empobrecido.
Se criaron ratas “listas” y ratas “torpes” en tres condiciones controladas diferentes:
- Condiciones normales.
- Ambiente enriquecido: jaulas más grandes con abundancia de objetos utilizables como juguetes.
- Ambiente empobrecido: jaulas especialmente pequeñas y sin ningún tipo de objeto.
Resultados:
- En condiciones normales, las diferencias entre ambos grupos resultaban evidentes (alta heredabilidad).
- En condiciones especiales (tanto enriquecidas como empobrecidas), dichas diferencias se hicieron inapreciables.
Estos resultados demuestran que puede darse una interacción genotipo/ambiente: en este caso, el ambiente enriquecido suple las carencias genéticas de las ratas «torpes»; por su lado, el ambiente empobrecido minimiza las ventajas genéticas para el aprendizaje de laberintos de las ratas «listas». Una conclusión interesante que podemos extraer puede resumirse diciendo que un alto valor de la heredabilidad no significa que ese rasgo no esté influido por factores ambientales.
AUTOEVALUACIÓN
GENÉTICA CUANTITATIVA DE LA CONDUCTA HUMANA
Con las herramientas de la genética cuantitativa, abordamos la cuestión de la base genética de algunos rasgos del comportamiento humano.
Inteligencia
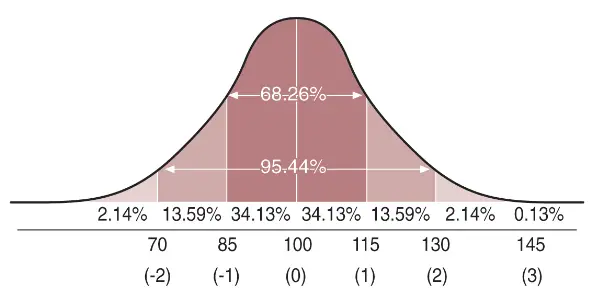
humana y la Campana de Gauss. La fundamentación teórica de
la distribución normal se puede encontrar en cualquier texto de
estadística.
Asumiendo que la inteligencia es un rasgo fenotípico de carácter cuantitativo, la medición de la inteligencia se realiza mediante tests psicológicos. Estos tests permiten clasificar a los individuos (aunque con un poco de practica, el más tonto puede obtener excelentes resultados). La distribución de la inteligencia en la población es «normal» y se adecua a una Campana de Gauss, lo que nos permite conocer qué puesto ocupa cada individuo en la población. p.ej., en los informes psicológicos que hacen los psicólogos escolares, los resultados suelen darse en percentiles, donde el valor del percentil señala únicamente qué porcentaje de individuos de la población están por debajo.
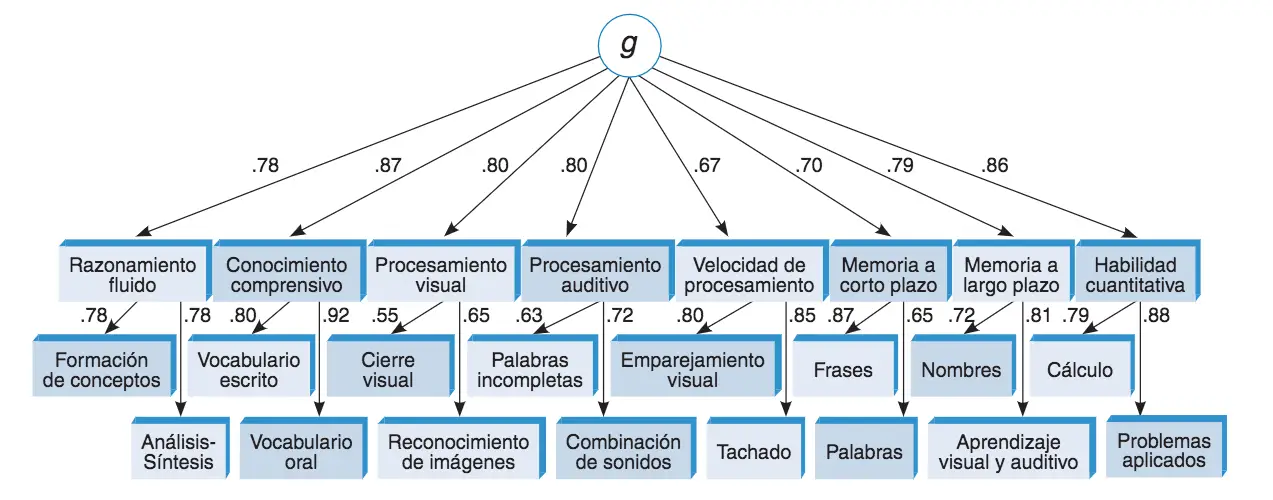
cognitivas con el llamado factor g.
- Podemos considerar que el CI es la medida del llamado factor g o factor de inteligencia general.
- Según las revisiones más modernas, la heredabilidad del CI evaluada a partir de estudios de gemelos monocigóticos criados por separado (MZS) se halla alrededor de 0.75.
- Los datos apuntan a que la influencia del ambiente compartido (el ser criados en un mismo ambiente) sobre el CI parece ser nula.
- Los factores ambientales que explican la variabilidad que no explica la variabilidad genética son peculiares para cada individuo y no, como se venía pensando, comunes para quienes viven en el mismo entorno familiar o social; algo similar puede decirse para los demás rasgos psicológicos.
- A medida que aumenta la edad, crece también el valor de la heredabilidad y disminuye la influencia del ambiente.
- Los valores de la heredabilidad en relación con el CI aumenten a medida que aumenta el estatus social (supuestamente un mayor estatus social va acompañado de más y mejores oportunidades de aprendizaje y experiencias; a más estatus, más enriquecimiento estimular del entorno).
Basándonos en el conjunto de datos existentes en la actualidad, podemos estar bastantes seguros que la mayor parte de la varianza genética de la inteligencia general o factor g es de tipo aditivo: los valores de la heredabilidad obtenido con gemelos no difiere de la obtenida a partir de la correlación entre padres e hijos.
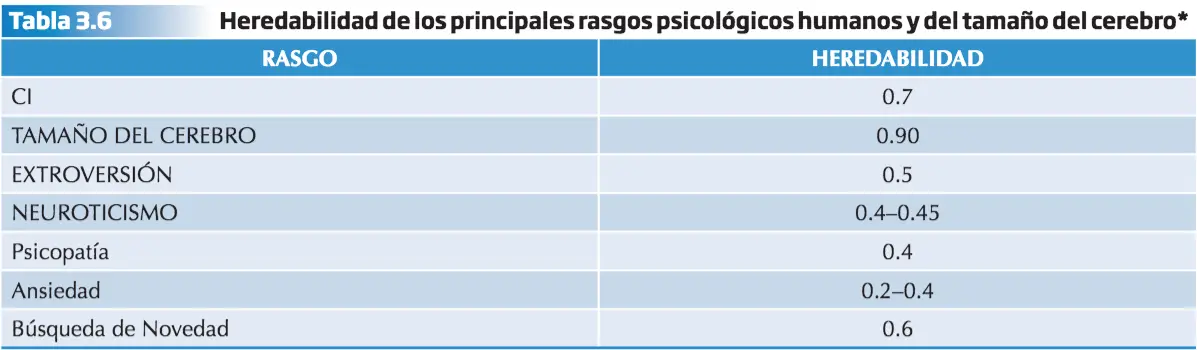
Psicopatología
Las enfermedades mentales más importantes tanto por su incidencia como por el deterioro que ocasionan en los pacientes son la esquizofrenia, las alteraciones del humor, con su doble vertiente, maníaca y depresiva, y los trastornos de ansiedad, de los que la neurosis obsesivo-compulsiva es el ejemplo más prominente.
La incidencia de la esquizofrenia es del 1 % en prácticamente todas las sociedades del mundo. El caso más curioso de concordancia genética gemelar (o porcentaje de gemelos que coinciden en padecer la enfermedad con respecto al total de pares de gemelos de los que uno de los dos la padece) de la esquizofrenia ha sido el de las cuatrillizas Genain (es nombre ficticio), quienes entre los 22 y los 24 años desarrollaron, todas ellas, síntomas esquizofrénicos de diversa gravedad.
Sobre las alteraciones del humor hay una cierta ambigüedad en la medida en que es difícil asegurar que la maniacodepresión o síndrome bipolar es distinto o sólo una forma más severa de la depresión unipolar y que ambas sean manifestaciones extremadas de los cambios de humor normales en cualquier ser humano, o bien enfermedades específicas. La incidencia del síndrome bipolar es del 1 %; por su parte el síndrome unipolar (sólo fase depresiva) es más frecuente, hasta un 5%; siendo el doble el número de mujeres que el de hombres que lo padecen. Entre un 15 y un 20% de estos pacientes se suicidan.
Entre los trastornos de ansiedad se encuentran la neurosis obsesivocompulsiva, los ataques de pánico y las fobias.
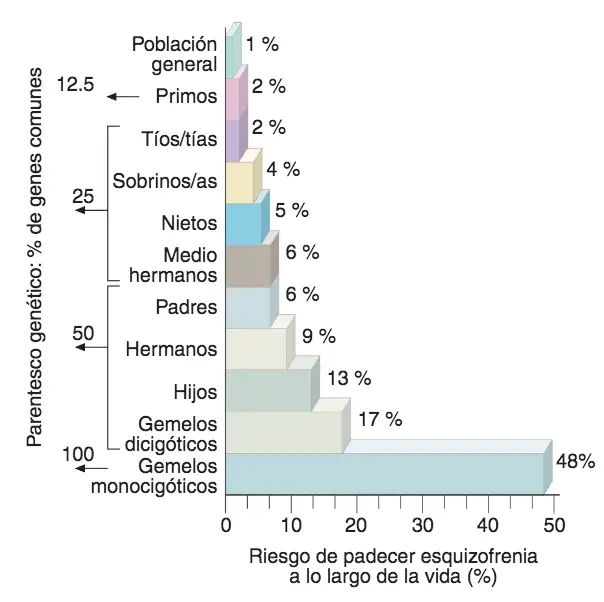
El riesgo familiar de esquizofrenia proporciona una evidencia de heredabilidad.
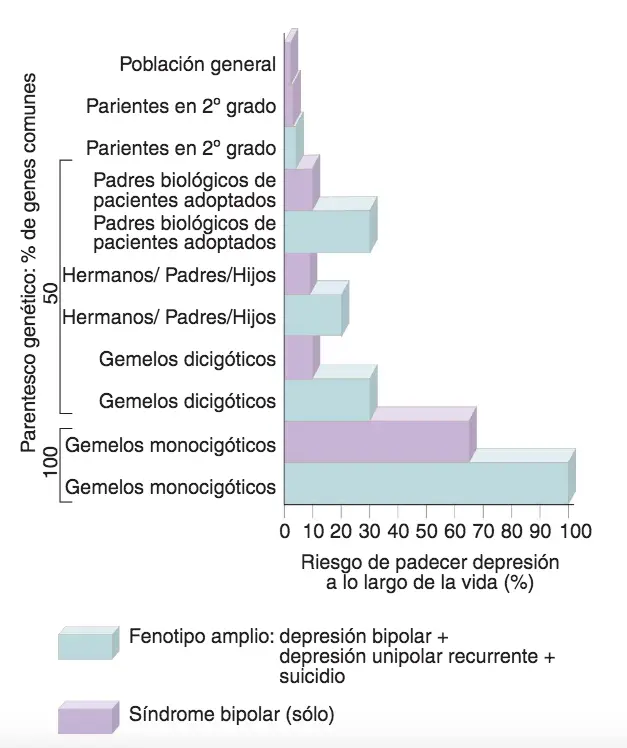
Riesgo de padecer síndrome bipolar o algún síntoma asociado a maniacodepresión entre diferentes grados de parentesco.
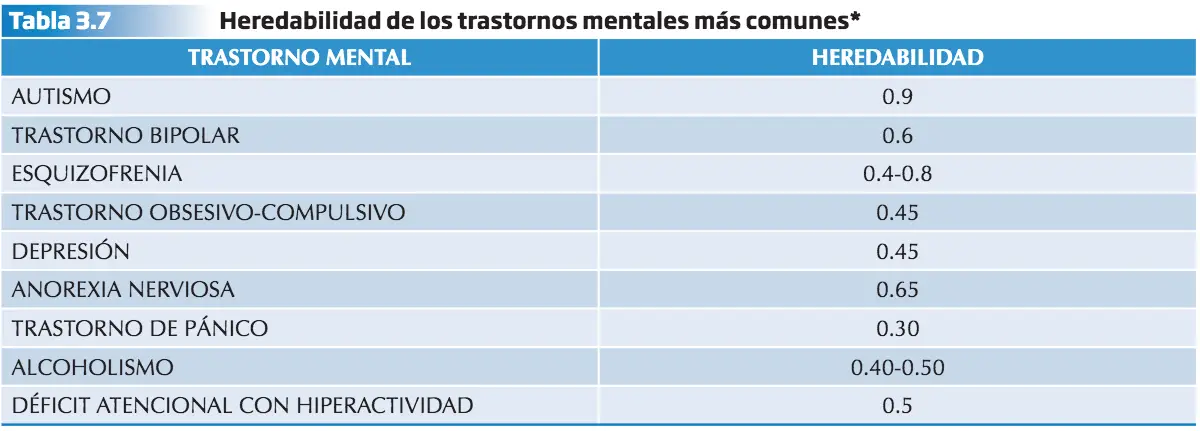
El valor que aquí se da, en realidad, es el de la heredabilidad del riesgo de padecer la enfermedad según el modelo de umbral de riesgo (liability threshold model)
Lección Magistral
AUTOEVALUACIÓN
REFERENCIAS
- Abril Alonso, A. (2016). Fundamentos de psicobiología (UNED (Sanz yTorres). Alcorcón (Madrid): Sanz y Torres.
- Powerpoint María Penado
- Apuntes Cari Blanco Rodríguez
- Apuntes Tania
- Profesora Mª Penado
- YouTube