

INTRODUCCIÓN
Aunque la idea de que la memoria era una facultad única dominó la historia del pensamiento científico durante siglos, a partir del siglo XIX, filósofos y psicólogos pensaban que la memoria no era una facultad o proceso único. Uno de ellos fue Maine de Biran (1804/1929) para el que existían 3 tipos de memoria: la memoria mecánica, la sensorial y la representativa. A finales del S. XIX, William James (1890), postulaba que la memoria se localizaba en el cerebro porque sin este la memoria no podía existir. En el siglo XX, McDougall (1923) señaló que había dos formas de reconocimiento: uno implícito y otro explícito; Bruner (1969) distinguió entre “memoria con registro” y “memoria sin registro”. Winograd (1975) diferenció entre conocimiento declarativo y conocimiento procedimental. Sin embargo, según Squire (2004), lo que faltaban eran estudios experimentales y no tratamientos o intuiciones psicológicas.
Resultados de estudios realizados con pacientes amnésicos. El estudio experimental sobre los sistemas de memoria comenzó con los estudios de Brenda Milner (1962), esta investigadora comprobó que el paciente H.M padecía amnesia anterógrada causada por una intervención, sin embargo, tras varios días de entrenamiento fue capaz de realizar tareas que requerían la coordinación mano-ojo. En esa época se consideraba que las habilidades motoras (como las demostradas por H.M) eran una forma de memoria de naturaleza inferior, menos cognitiva que otros tipos de memoria, y que lo que se denominaba habitualmente memoria estaba deteriorado en este paciente. Sin embargo, otros estudios realizados pocos años después también con pacientes amnésicos (Warrington y Weiskrantz, 1970; 1978) mostraron la buena actuación de estos en otras tareas distintas de las tareas motoras utilizadas por Brenda Milner.
El hecho de que los pacientes amnésicos actuaran normalmente en tareas de memoria en las que no se mencionaba la necesidad de recuperar voluntariamente la información prueba que el fenómeno del priming es una forma de memoria diferente de la que se encuentra deteriorada en los pacientes amnésicos (Schacter y Buckner, 1998; Tulving y Schacter, 1990). La mayor evidencia de la separación entre estas dos formas de memoria, la que se encuentra deteriorada en la amnesia y la que se mantiene en estos enfermos proviene de otros estudios realizados por Squire con enfermos amnésicos que mostraban priming intacto mientras actuaban al azar con los mismos estímulos en pruebas de reconocimiento en las que les pedía que recuperaran la información voluntariamente e indicaran si se trataba de un estímulo presentado previamente (antiguo) o si se trataba de un estímulo nuevo (Levy, et al., 2004).
Investigaciones realizadas con animales en laboratorio permitieron proponer una clasificación de la memoria más precisa (Mishkin, 1982). Zola-Morgan y Squire (1984) utilizaron monos como sujetos experimentales. El objetivo era comprobar qué estructuras cerebrales lesionadas resultaban más importantes para la actuación normal de la memoria. Utilizaron la conocida como “tarea retrasada de no igualación a la muestra” que consistía en mostrar al mono un objeto y esconder debajo del mismo una recompensa. El mono levantaba el objeto y obtenía la recompensa. Después de varios retrasos, se presentaban al mono dos objetos. Uno de esos objetos era el mismo presentado anteriormente. El otro, era un objeto nuevo debajo del cual estaba oculta la recompensa. El animal tenía que recordar el objeto que había levantado antes ya que la recompensa se encontraba ahora debajo del objeto nuevo.
Los resultados más importantes de las investigaciones con animales a los que se les producían lesiones en el cerebro fueron que la actuación se deterioraba en función del tiempo que mediaba entre la presentación del objeto único y la presentación de los dos objetos (el “antiguo” y el “nuevo”). En estos estudios los investigadores lesionaron distintas zonas del cerebro y los resultados mostraron que la lesión de la amígdala no desempeñaba un papel importante en la pérdida de memoria conocida como síndrome amnésico. En cambio, la lesión de áreas corticales que rodeaban al hipocampo y la del mismo hipocampo, desempeñaban un papel primordial para la realización de la tarea de memoria. Demostraron también que podía producirse una amnesia severa, aunque no se lesionara el hipocampo sino otras zonas próximas a él como las cortezas parahipocámpicas y perirrinales.
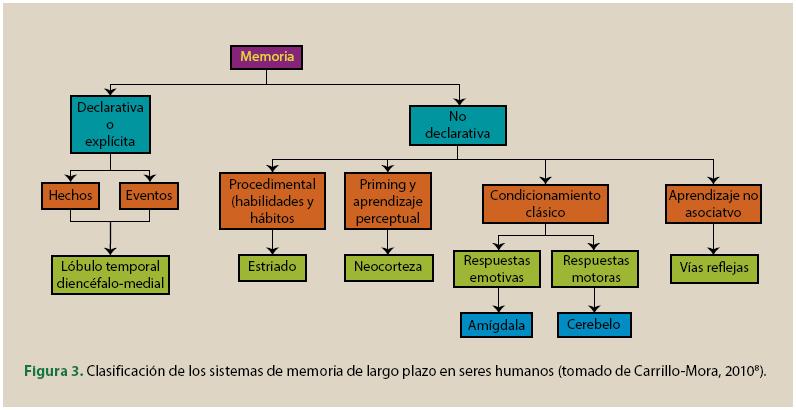
A finales del siglo XX algunos psicólogos ya pensaban que existían varios sistemas de memoria (Schacter y Tulving, 1994; Tulving, 1985; Squire y Zola-Morgan, 1988). La memoria declarativa corresponde a uno de esos sistemas. Esta memoria es la que se deteriora en la amnesia y está ligada a la lesión del lóbulo temporal medio y el diencéfalo. La memoria declarativa permite recobrar material y apoya la codificación de los recuerdos por sus relaciones establecidas entre múltiples hechos y elementos. Además, es representacional y constituye una forma de modelar el mundo externo porque puede ser verdadera o falsa. La memoria declarativa se divide en memoria semántica que es un tipo de memoria sobre hechos, y memoria episódica que consiste en la recuperación de un hecho o evento en el contexto en el que dicho evento ocurrió. El sistema de memoria episódica requiere, además de la participación del sistema temporal medio diencefálico, la participación de los lóbulos frontales (Shimamura y Squire, 1987).
El hipocampo y otras estructuras relacionadas son fundamentales para la formación, la reorganización y consolidación de la memoria de larga duración. Nuevas líneas de trabajo sugieren que el lóbulo temporal medial desempeña un papel fundamental en el proceso de almacenamiento de la información en la memoria. La lesión de estas estructuras conserva la memoria remota y deteriora memorias más recientes (Restivo, Vetere, Bontempi y Ammassari-Teule, 2009; Squire, 2009b). Al contrario de la memoria declarativa, la memoria no declarativa se expresa a través de la actuación y no puede ser ni verdadera ni falsa. Estas memorias se muestran mediante la reactivación de los sistemas en los que originalmente se produjo el aprendizaje.
¿QUÉ SON LOS SISTEMAS DE MEMORIA?
La propuesta de la existencia de distintos sistemas de memoria se debe a Tulving et al. y sus colegas, para los que un sistema es una estructura anatómica y evolutivamente distinta de otros sistemas de memoria y se diferencia por sus métodos de adquisición, sistema de representación y recuperación del conocimiento (Tulving, 1983, 1985). Además, se desarrolla a ritmos distintos y se deteriora de forma diferente debido a la enfermedad y al envejecimiento (Schacter y Tulving, 1994).
Los distintos sistemas de memoria existentes en el cerebro funcionan en paralelo para soportar y mantener la conducta del individuo. Un suceso negativo de la infancia puede producir un recuerdo declarativo estable relacionado con ese evento, pero también puede producir miedo duradero no declarativo.
Criterios para determinar la existencia de un sistema de memoria
Schacter y Tulving (1994) propusieron tres criterios para identificar un sistema de memoria: Operaciones de inclusión de clase; Propiedades y relaciones; Disociaciones convergentes.
• Operaciones de inclusión de clase. Un sistema de memoria permite realizar una amplia variedad de tareas de una determinada clase. Por ejemplo, el sistema de memoria de trabajo permite mantener durante un tiempo breve un número pequeño de elementos de una cierta clase en un almacén y realizar operaciones cognitivas con ese material. El sistema de memoria episódica permite recordar acontecimientos de la vida pasada, aunque el tiempo transcurrido sea muy largo. Esto no es posible dentro de la MCP. Cuando se produce un accidente, una enfermedad o un trauma que hace que una clase de funciones de memoria se altere o se elimine, puede ser la consecuencia de los cambios en la forma de operar de ese sistema. Las regiones cerebrales implicadas son los componentes neurales del sistema. Los cambios cerebrales pueden producirse por causas muy diversas, incluidas la privación de sueño, trauma emocional, hipnosis, ingestión de medicamentos y uso de drogas, enfermedades, lesiones producidas quirúrgicamente, etc.
• Propiedades y relaciones. un sistema de memoria debe poder describirse mediante una lista de propiedades que definen su identidad y sus relaciones con otros sistemas. Estas propiedades deben incluir sus reglas de funcionamiento, el tipo de información que contiene y su substrato neural. Un sistema debe servir para algo.
• Disociaciones convergentes. Los sistemas de memoria deben mostrar la existencia de una serie de disociaciones convergentes que incluyan las observadas en diferentes tipos de tareas de memoria, diferentes poblaciones y las obtenidas utilizando diferentes técnicas (Schacter, 1992; Tulving, 1983).
Los cinco sistemas de memoria
Utilizando los criterios enumerados se han identificado 5 sistemas de memoria (Schacter y Tulving, 1994; Schacter, Wagner, y Buckner, 2000):
- • El sistema de memoria de trabajo.
- • El sistema ele memoria procedimental
- • El sistema de representación perceptual
- • El sistema de memoria semántica
- • El sistema de memoria episódica
Sistema de memoria de trabajo. es el encargado del mantenimiento y almacenamiento temporal de la información. Aunque existen diferentes versiones el mejor conocido es el de Baddeley (1986, 1994). Está formado por un ejecutivo central que es un mecanismo de control atencional que comprende dos sistemas subsidiarios. El bucle articulatorio, especializado en el procesamiento de la información verbal, y la agenda visoespacial, que se ocupa de la información visoespacial y de la adquirida por el tacto. En la última especificación, Baddeley incluyó también un retén o buffer episódico. La información permanece activa durante un periodo de tiempo breve y su capacidad es limitada. Al cabo de unos segundos, la información que no se repite, decae y desaparece. Las pruebas para evaluar este sistema contienen un pequeño número de elementos y a continuación se presenta el material que se debe manipular.
Sistema de la memoria procedimental. Desde un punto de vista evolutivo, es el más antiguo y menos explorado. Está relacionado con la actuación y la acción y la adquisición de sus contenidos es gradual. Está relacionado con “saber cómo” en vez de estarlo con “saber qué” (Tulving, 1983, 1985). Por ejemplo, saber que “los trabajadores tenemos derecho vacaciones pagadas por la empresa” o que “la educación pública es un bien social”, depende de la memoria declarativa. Por el contrario, saber ejercer los derechos laborales, o denunciar un delito contra los derechos de los trabajadores depende de la memoria procedimental. Para poder aprender este procedimiento, la persona tiene que realizar una serie de acciones. Escuchar simplemente una serie de instrucciones, o mirar como otras personas lo hacen, no es suficiente para aprender y demostrar que se posee esta habilidad. Las pruebas que se utilizan para evaluar este tipo de memoria son indirectas y no exigen que la persona tenga que recuperar conscientemente el episodio original en el que se produjo el aprendizaje. Este tipo de memoria es muy resistente al olvido que se produce por la falta de práctica, pero la habilidad se vuelve a adquirir rápidamente con la práctica.
Sistema de representación perceptual. Se trata de un sistema no declarativo que está formado de una serie de módulos específicos que operan sobre la información sensorial relacionada con la forma y la estructura de las palabras y los objetos. La información en el sistema se registra en un formato específico. Los datos sobre los que se basa la propuesta de la existencia de este sistema provienen de estudios del priming. El priming se muestra porque cuando tenemos que responder al mismo estímulo en una segunda ocasión, o respondemos a un estímulo que es similar físicamente o que tiene el mismo sentido, la respuesta es más rápida y/o más precisa que lo fue cuando respondimos dese estímulo la primera vez. Lo que tienen en común las tareas para evaluar este sistema es que en ellas no se pide la recuperación consciente de la información. Las variables que influyen en el priming son diferentes a las que influyen en los sistemas de la memoria declarativa. Este sistema está preservado en pacientes amnésicos y el Alzheimer.
Sistema de memoria episódica. Este sistema contiene recuerdos de experiencias personales sobre acontecimientos y episodios. La persona tiene que retroceder mentalmente en el tiempo para examinar su pasado (Tulving, 2002). La información del contexto es fundamental en este sistema. Se cree que tiene una capacidad ilimitada y el olvido se produce por interferencia. Se trata de la forma de memoria superior y se desarrolla en una escala temporal diferente a la de otros sistemas. El sistema puede lesionarse sin que otros sistemas de memoria se destruyan. Los contenidos de este sistema de memoria se ponen a prueba mediante pruebas de reconocimiento y de recuerdo, libre y señalado de la información presentada previamente.
Sistema de memoria semántica. Se trata de un sistema de memoria declarativo, sus contenidos deben traerse voluntariamente a la consciencia. El sistema incluye el conocimiento general, conocimiento de hechos, conceptos y conocimiento del lenguaje (Tulving, 1983). Se caracteriza por combinar el conocimiento consciente sobre su contenido con la falta de consciencia sobre el episodio de aprendizaje. Se cree que tiene una capacidad ilimitada y que permite mantener la información durante un tiempo ilimitado. El olvido se produce por no poder acceder temporalmente a la información porque se produce interferencia. Se trata más bien de no poder acceder a la información temporalmente más que no poder acceder a ella de manera permanente. Esta memoria se evalúa a través de preguntas de conocimiento general, la realización de tareas de formación de categorías o tareas similares.
¿SISTEMAS O PROCESOS DE MEMORIA?
El enfoque de los sistemas de memoria ha sido criticado por los defensores de las teorías del procesamiento (Roediger, Buckner y McDermott, 1999). Las principales críticas se refieren: A la falta de criterios unánimemente aceptados para generar los cinco sistemas de memoria diferentes; a que la propuesta se centra sobre todo en la existencia de disociaciones, porque cualquier diferencia en un sentido en o en otro puede tomarse como evidencia para apoyar la distinción entre sistemas; y en que tiene dificultades para explicar los resultados observados al principio y al final del ciclo vital. (Surprenant y Neath, 2009)
La mayor fortaleza de los sistemas de memoria está en su capacidad para explicar los resultados obtenidos de los pacientes amnésicos que tienen dificultades para realizar tareas de memoria episódica, pero que realizan normalmente tareas de memoria semántica y memoria procedimental (Gabrieli, 1999; Parkin, 1999). Los defensores de las teorías de procesamiento se basan en los procesos de codificación y recuperación en lugar de postular sistemas de memoria y su posible localización cerebral en la que se almacena la memoria (se basan en el tipo de procesamiento que la persona pone en marcha). La memoria para ellos es un proceso activo y constructivo (postura de Bartlett). La recuperación de la información a partir de la memoria dependerá de las claves ambientales y de la información procesada previamente por el individuo (Surprenanl y Neath, 2009).
Craik y Lockhart (1972) ofrecieron la primera propuesta moderna que sugería que el procesamiento del individuo era más importante que el almacén en el que se recogía. Propusieron que la memoria es el resultado de una serie sucesiva de análisis que iba desde un análisis superficial (rasgos físicos), hasta un nivel profundo (significado e información semántica). Supusieron que, a mayor nivel de procesamiento, el recuerdo es mejor. La variable fundamental era la profundidad de procesamiento mientras que el número de repeticiones mentales y el tiempo desde la codificación inicial no tenían interés. El principal problema de los niveles de procesamiento es su falta de objetividad, ya que no es posible “a priori” evaluar la profundidad de un tipo particular de procesamiento.
La propuesta de los defensores de la transferencia de procesamiento apropiado sugiere que un tipo de procesamiento durante la fase de estudio producirá mejor actuación si resulta apropiado para el tipo de prueba que se utilice para evaluar la memoria. El procesamiento que se realice durante la fase de codificación y el que se realice durante la recuperación de la información son muy importantes. Según Roediger y Blaxton (Blaxton, 1989; Roediger y Blaxton, 1987), la mayor parte de los estudios implícitos y explícitos contienen una confusión. Normalmente, las tareas de memoria indirectas se basan en el procesamiento guiado por los datos, de tipo perceptivo o de nivel más bajo (indicar si una palabra está en mayúsculas o en minúsculas). En el caso de las tareas directas lo que se destaca son los aspectos semánticos o de más alto nivel, se basan en el procesamiento guiado conceptualmente. Por ejemplo, si las palabras tienen una connotación agradable o desagradable, o si una palabra es un sinónimo de otra palabra. Blaxton (1989) concluyó que lo importante era el tipo de procesamiento del sujeto y no el sistema de memoria subyacente.
Una dificultad a la que se enfrentan las teorías de procesamiento es cómo definir y especificar con claridad y objetivamente el tipo de procesamiento que pone en marcha el individuo cuando realiza una tarea de memoria. Tanto las teorías de la profundidad de procesamiento como las teorías de la transferencia de procesamiento apropiado carecen de una especificación formal. La mayor parte de las tareas exigen ambos tipos de procesamiento y es difícil, por no decir imposible, evaluar la contribución relativa de cada tipo de procesamiento.
Otra dificultad añadida es la posibilidad de probar su falsedad porque cuando el resultado de un estudio sale en la dirección esperada, se interpreta que es porque los individuos están usando el procesamiento apropiado. Pero, cuando los resultados no son los esperados, estos se explican porque algunos están utilizando un tipo de procesamiento diferente. A pesar de todo esto, la mayor debilidad de esta postura es la dificultad para explicar los datos que muestran una destrucción total de la memoria episódica en la amnesia mientras se encuentra conservada la memoria semántica y la memoria implícita. Esta es, precisamente, la mayor fortaleza de la teoría de los sistemas de memoria.
La idea de la existencia de distintos sistemas cerebrales implicados en las distintas memorias parece en la actualidad, más consistente con los datos psicológicos, biológicos y de imágenes cerebrales disponibles (Squire, 1994; 2004).
SISTEMAS DE MEMORIA Y CEREBRO HUMANO
El concepto de sistemas de memoria es uno de los constructos fundamentales de la neurociencia cognitiva. La idea de que la memoria estaba formada por un número de componentes funcionales, neuralmente diferentes, se basó en las disociaciones encontradas en la actuación de pacientes neurológicos en distintos tipos de tareas de memoria. Esta idea está respaldada por los resultados de un número cada vez mayor de estudios que han utilizado neuroimágenes funcionales (Cabeza y Kingstone, 2005).
La memoria declarativa depende del sistema temporal medial diencefálico. Esta zona del cerebro y el hipocampo se encuentran lesionados en la amnesia, razón por la que los amnésicos actúan mal en pruebas de memoria episódica mientras otros tipos de memoria se encuentran preservados. Esta zona es también la primera en deteriorarse en la demencia tipo Alzheimer. No es casualidad que uno de los primeros signos de la enfermedad son los trastornos de la memoria episódica.
La investigación con animales ha mostrado que el cerebelo es esencial para el condicionamiento del parpadeo (McCormick, Clark, Lavond, y Thompson, 1982). Este tipo de aprendizaje se encuentra preservado en animales con lesiones en el hipocampo (Schmaltz y Theios, 1972) y en pacientes amnésicos (Clark y Squire, 1998). También se ha encontrado que de la amígdala depende el condicionamiento de respuestas emocionales (LeDoux, 2004).
APORTACIONES A PARTIR DE LOS ESTUDIOS CON IMÁGENES CEREBRALES
La neurociencia cognitiva, ha hecho posible estudiar cómo la función cerebral soporta las actividades mentales. El número de artículos y la inversión económica para financiar centros de investigación a nivel mundial aumenta de manera exponencial han aumentado.
Los orígenes de la resonancia magnética funcional (fMRI) hay que buscarlos en la resonancia magnética nuclear, sin embargo, para que las imágenes resulten útiles, es necesario disponer de un marcador. En 1990 se identificó este marcador al descubrirse que la intensidad de la señal de algunas imágenes de resonancia disminuía en presencia de sangre paramagnética deoxigenada. Lo que hoy se conoce como contraste BOLD (siglas del término inglés blood-oxigenation-level-dependent) constituye la base de los estudios de resonancia magnética funcional (fMRI) y su utilización se basa en las propiedades magnéticas de la hemoglobina que funciona como un elemento de contraste endógeno. Así, se pueden detectar los cambios producidos en el flujo sanguíneo cerebral durante una tarea cognitiva.
Un escáner normal tiene campos magnéticos de 1,5 y 3 Teslas (los modernos hasta 7). La fMRI utiliza el escáner para registrar los cambios en el metabolismo cerebral asociados a la actividad neuronal que se produce en distintas zonas del cerebro mientras se realizan distintas tareas cognitivas. La señal fMRI se registra a partir de los protones de hidrógeno existentes en las moléculas de agua. Las imágenes por fMRI podían utilizarse para hacer mapas de la función hemodinámica del cerebro, y su correspondencia con las operaciones mentales.
Las principales ventajas de la resonancia magnética funcional (técnica más utilizada en la actualidad) son:
- No es necesario inyectar isótopos radiactivos en los pacientes (es una técnica inocua).
- Su resolución espacial (alrededor de 1,5 x 1,5 mm) es superior a otras técnicas de imágenes.
- El tiempo para realizar el escaneo es corto (1,5 a 2 segundos).
Sin embargo, es una técnica difícil y cara, el diseño de los experimentos es complicado y el análisis de datos complejo, por eso la investigación en este campo requiere de la colaboración multidisciplinar. Aun así, tiene ventajas con respecto a la tomografía de emisión de positrones (PET), que requiere la inyección de isótopos radiactivos, múltiples adquisiciones de imágenes y más tiempo de registro.
IMPORTANCIA DE LAS NEUROIMÁGENES FUNCIONALES EN LA INVESTIGACIÓN COGNITIVA
William James (1890), trató de las relaciones existentes entre conducta y cerebro. Sin embargo,fuePavlovel que tuvo una visión de estas relaciones y que en su época parecía fantástica. La visión fue: “Si a través del cráneo pudiéramos ver el cerebro de una persona mientras está pensando conscientemente, y si el lugar de máxima excitación fuera luminoso, veríamos en la superficie cerebral una zona brillante con bordes fantásticos fluctuando constantemente en tamaño y forma, rodeados de una oscuridad más o menos profundacubriendo el resto de los hemisferios”.
El descubrimiento de las técnicas de imágenes cerebrales en los 70(la tomografía de emisión de positrones -PET, y después las imágenes por resonancia magnética-MRI), permitió hacer realidad el sueño de Pavlov. Estas permiten observar (cada vez con más detalle) el funcionamiento del cerebro y mapear los cambios locales de la circulación y elmetabolismo cerebral asociados con los cambios en la actividad celular del cerebro(Posner y Raichle).1994).
Imágenes funcionales en la investigación sobre la memoria de trabajo
La memoria de trabajo sirve para la retención temporal y la manipulación de la información que acaba de presentarse y que ya no está presente en el medio ambiente. Las representaciones de este sistema de memoria tienen una corta duración que puede prolongarse mediante su mantenimiento activo (Baddeley, 1986; D ‘Esposito, 2007).
Estudios con monos en los que se registraba la actividad de las neuronas de la corteza prefrontal lateral (PFC) mostraron la existencia de niveles sostenidos de actividad neuronal durante un intervalo de retención breve (Fuster y Alexander, 1971). Esta actividad es el nexo de unión entre la clave proporcionada por el estímulo y la respuesta contingente del individuo. Estudios más recientes realizados con humanos han apoyado estos resultados, lo que ha hecho que en la actualidad exista consenso en que la corteza prefrontal lateral (PFC) es la zona del cerebro implicada en la realización de distintas tareas de memoria de trabajo, es decir, es esencial para el mantenimiento de las representaciones internas de la información durante un tiempo breve. Distintos tipos de tareas de memoria de trabajo producen diferentes patrones de activación en la corteza prefrontal. Por ejemplo, estímulos de tipo verbal activan el hemisferio izquierdo, mientras que estímulos de tipo espacial activan el hemisferio derecho.
CONTRIBUCIÓN DE LAS ÁREAS PREFRONTALES A LA MEMORIA DE TRABAJO: EVIDENCIA A PARTIR DE UN ESTUDIO DE FMRI
Objetivo. Investigar la disociación entre los procesos necesarios para el mantenimiento de la información en la memoria de trabajo y el proceso implicado en la manipulación de dicha información.
Paradigma experimental.Se utilizó una tarea de respuesta retrasada en la que un conjunto de 5 letras se presentaron simultáneamente en un orden aleatorio, seguido inmediatamente de la presentación de la instrucción:“HACIA ADELANTE” o “ALFABETIZAR”, seguida de una cruz de fijación que estaba presente en el ordenador 8 segundos. A continuación, aparecía una señal que indicaba responder apretando un botón.
Los participantes realizaron dos tipos de ensayos en un orden aleatorio dentro del escáner para registrar la actividad cerebral:
- Mantener en su memoria una secuencia de 5 letras presentadas en un orden aleatorio durante un retraso.
- Manipular una secuencia de 5 letras colocando las letras por orden alfabético durante el periodo de retraso.
Resultados.Durante el periodo de retraso encontraron en ambos tipos de ensayos, activación en las cortezas prefrontal dorsolateral y ventrolateral. La actividad dorsolateral fue mayor en los ensayos de manipulación en la memoria de trabajo.
Conclusiones.Los estudios sugieren que la manipulación de la información en la memoria de trabajo está asociada con una actividad cerebral más extensa que el simple mantenimiento de la información. Además, la diferencia entre manipulación y mantenimiento es mayor en la corteza prefrontal dorsal.
En resumen, la corteza prefrontal (PFC) parece que es la región cerebral más importante para la memoria de trabajo. Los resultados de muchos estudios sobre la memoria de trabajo realizados con fMRI han mostrado que la corteza prefrontal es esencial para el mantenimiento activo de las representaciones internas de la información durante un tiempo breve. La memoria de trabajo parece que no está localizada en una única zona cerebral, sino que emerge de la interacción entre la corteza prefrontal y otras regiones parietales posteriores (Curtis y D ‘ Esposito, 2003; D ‘Esposito, 2007; Pudas et al., 2009).
Imágenes funcionales y memoria episódica
Estudios con técnicas de imágenes cerebrales han permitido estudiar los patrones de activación cerebral y el funcionamiento del hipocampo y de la corteza temporal media (MTL) durante la realización de tareas de memoria episódica y han mostrado que otras regiones (cortezas prefrontal y parietal) también influyen. Gracias a la utilización de fMRI se ha encontrado que la activación del lóbulo temporal medio es importante para la formación de la memoria episódica. Esta zona está muy interconectada con otras áreas de asociación que rodean al hipocampo como son las cortezas perirrinal, entorrinal y parahipocampal que tienen proyecciones en el hipocampo. Estos estudios han mostrado que la activación del hipocampo, de la zona perirrinal y del giro posterior parahipocampal correlaciona con tareas de memoria episódica (Davachi y Dobbins, 2008).
En un estudio, los investigadores presentaron a los participantes dentro del escáner una lista de adjetivos para que los estudiaran (Davachi, Mitchell y Wagner, 2003) mientras registraban las imágenes cerebrales. Antes de la presentación de cada palabra se presentaba una señal para que realizaran con ella una de las dos tareas de codificación (imaginar o leer). Evaluaron la memoria episódica con una prueba que permitía:
- Identificar los ítems que los participantes olvidaron pero que habían estudiado previamente.
- Identificar los ítems que los participantes reconocían como presentados, pero no podían recordar el contexto (tarea de codificación) en el que habían sido presentados (reconocimiento del ítem).
- Reconocían el ítem y recordaban el contexto en el que había sido presentado (un índice de reconocimiento del ítem más recuperación episódica).
Los resultados de las imágenes cerebrales mostraron que la activación en el hipocampo durante la codificación correlacionaba con la recuperación episódica de los detalles que rodeaban los eventos, primeramente, o después la memoria relacional, pero no diferenciaba entre los ítems reconocidos y los olvidados. Este patrón de activación no se encontró en todo el lóbulo temporal medio. La activación en la zona adyacente de la corteza perirrinal correlaciona con el reconocimiento del ítem independientemente de la recuperación de los detalles episódicos.
El reconocimiento de la experiencia previa con los estímulos depende de dos procesos:
- Recuperación del estímulo con el contexto de otra información asociada con la experiencia.
- Familiaridad con los rasgos del estímulo.
Estudios con animales sugieren que estos dos procesos (recuperación y familiaridad), dependen de subregiones del lóbulo temporal medio:
- El hipocampo desempeña un papel importante en la recuperación, pero no la familiaridad.
- La región parahipocampal contribuye a la recuperación a través de la recuperación de la información del contexto espacial, mientras que la familiaridad depende de la corteza perirrinal.
En resumen, el hipocampo es importante para la codificación de relaciones y para la recuperación de la información sobre relaciones. La corteza perirrinal interviene en la codificación del ítem y en el reconocimiento del mismo, independientemente del contexto.
Correlatos neurales de la memoria implícita
Cuando se repiten los estímulos, la actividad neural asociada con el procesamiento de estos estímulos disminuye en un amplio número de áreas cerebrales comparado con la actividad mostrada por los estímulos presentados por primera vez (Henson, 2003). Esta reducción se ha denominado de varias maneras: priming neural, supresión con la repetición y adaptación. Aunque el resultado habitual es la reducción, se ha encontrado aumento de la actividad cuando los estímulos son no familiares (James et al., 2002).
El priming conductual implica varios procesos, puede tener efectos en el procesamiento perceptivo posterior del estímulo (priming perceptivo) y puede producir procesamiento amodal o semántico, que no depende de la modalidad a la que se presenten los estímulos. Estudios con resonancia magnética funcional en jóvenes han mostrado la sensibilidad de las regiones visuales (occipito-temporales) posteriores de la corteza cerebral a manipulaciones perceptivas de los estímulos que implican el cambio de alguna dimensión estimular de la fase de estudio a la fase de prueba, y que el priming se mantiene a través de transformaciones de las características físicas del objeto (color, punto de vista, tamaño o ejemplar), e incluso con el cambio de modalidad (visión-tacto, tacto-visión).
Estudios realizados con imágenes cerebrales han mostrado que las zonas cerebrales implicadas en procesos de memoria implícita y explícita pueden disociarse porque las áreas corticales activas son diferentes y no se solapan con las regiones cerebrales que predicen el priming cuando se repiten los mismos estímulos (Schott, et al., 2006). La actividad cerebral que predice priming difiere de la que predice memoria explícita. La actividad relacionada con el priming se produce en las áreas de la vía visual ventral (corteza extriada, giro frontal inferior, ambas bilateralmente, y giro fusiforme izquierdo). Estas zonas cerebrales están implicadas en la percepción e identificación de los estímulos. Los mecanismos perceptivos y de identificación del objeto operan durante el primer encuentro con el estímulo que después, con la repetición va a mostrar los efectos del priming (Tulving y Schacter, 1990). Resultados como los de Schott y cols. cuestionan las teorías que afirman que la memoria implícita y explícita es una única memoria que lo que hacen es reflejar modos de acceso diferentes a la hora de la recuperación a los mismos trazos de memoria episódica.
Por otro lado, otros estudios de procesamiento conceptual o semántico con tareas de clasificación semántica, han mostrado que la facilitación con la repetición depende de las regiones temporales y frontales. Cambios en zonas del giro frontal inferior y del giro temporal medio e inferior se han relacionado con el aumento de la fluencia en la recuperación de la información semántica que se produce con la repetición (Buckner et al., 2000; Wiget al., 2005). Estos dos tipos de procesos se cree que son diferentes de los procesos guiados perceptivamente (priming perceptivo), y reflejen procesamiento abstracto de atributos léxicos o semánticos.
Schacter y cols. (2007) revisaron estudios y sugirieron que la reducción de la actividad cortical durante el priming supone la existencia de dos mecanismos diferentes:
- Uno se refiere a los efectos de especificidad estimular que para los estímulos visuales parecen ser más pronunciados en las regiones visuales primarias, con el priming categorial ocurriendo en regiones más anteriores (fusiforme y áreas parahipocámpicas), con las regiones frontales e inferotemporales sensibles a los rasgos abstractos y conceptuales de los estímulos.
- Los estudios que han comprobado las correlaciones entre priming conductual y neural sugieren que estas correlaciones son significativas en las regiones prefrontales a través de muchas tareas y diferentes procedimientos, mientras que el priming neural apenas está correlacionado con el priming conductual en las regiones cerebrales posteriores.
En resumen, las disociaciones encontradas en pacientes neurológicos (amnesia y demencia), los resultados de investigaciones con animales lesionados y estudios con técnicas de imágenes cerebrales apoyan la existencia de distintos sistemas de memoria en el cerebro humano.
RESUMEN
La memoria no era una facultad o proceso único. A finales del siglo XIX William James en 1890, la memoria se localizaba en el cerebro porque sin éste, la memoria no podía existir. En el siglo XX, Mc Dougall, dos formas de reconocimiento, uno implícito y otro explícito; Bruner “memoria de registro” y “memoria sin registro”
Resultados de estudios realizados con pacientes amnésicos. Comenzó con los estudios neuropsicológicos de Brenda Milner con el paciente H.M
El hecho de que los pacientes amnésicos actuaran normalmente en tareas de memoria en las que no se mencionaba la necesidad de recuperar voluntariamente la información prueba que el fenómeno del priming es una forma de memoria diferente de la que se encuentra deteriorada en los pacientes amnésicos.
Investigaciones realizadas con animales. Los estudios de laboratorio realizados con animales permitieron proponer una clasificación de la memoria más precisa utilizaron monos como sujetos experimentales. El objetivo de sus estudios fue comprobar qué estructuras cerebrales lesionadas resultaban más importantes para la actuación normal de la memoria, utilizaron una tarea conocida como “tarea retrasada o de no igualación a la muestra”, consistía en mostrar al mono un objeto y esconder debajo del mismo una recompensa. El mono levantaba el objeto y obtenía la recompensa. Después de varios retrasos, se presentaban al mono dos objetos. Uno de estos objetos era el mismo presentado anteriormente. El otro, era un objeto nuevo debajo del cual estaba oculta la recompensa. El animal tenía que recordar el objeto. La recompensa se colocaba debajo del objeto de nuevo, el mono seguía perseverando en la respuesta que antes había tenido.
Los resultados más importantes de las investigaciones realizadas con animales a los que se les producían lesiones en ciertas zonas del cerebro y se comprobaba su función del tiempo que mediaba entre la presentación del objeto único y la presentación de los dos objetos. Los resultados mostraron que la lesión de la amígdala no desempeñaba un papel importante en la pérdida de memoria que se conocía como síndrome amnésico. Sin embargo, la lesión de las áreas corticales que rodeaban al hipocampo y la del mismo hipocampo, desempeñaba un papel primordial para la realización de la tarea de memoria. La memoria declarativa se divide en memoria semántica que es un tipo de memoria sobre hechos y memoria episódica que consiste en la recuperación de un hecho o evento en el contexto en el que dicho evento ocurrió. Este sistema de memoria episódica requiere además de la participación del sistema temporal medio diencefálico, la participación de los lóbulos frontales.
El hipocampo y otras estructuras relacionadas son fundamentales para la formación, la reorganización y la consolidación de la memoria de larga duración. En la actualidad nuevas líneas de trabajo sugieren que el lóbulo temporal medial desempeña un papel fundamental en el proceso de almacenamiento de la información en la memoria.
¿Qué son los sistemas de memoria? Tulving y sus colegas, para los que un sistema es una estructura anatómica y evolutivamente distinta de otros sistemas de memoria y se diferencia por sus métodos de adquisición, sistema de representación y recuperación del conocimiento, se desarrolla a ritmos distintos y se deteriora de forma diferente debido a la enfermedad y el envejecimiento.
Los cinco sistemas de memoria. El primero, más grande y el menos explorado es el sistema de la memoria procedimental, relacionado con la actuación y la acción. La adquisición de sus contenidos es gradual, se dividió distinguiendo entre acción (aprendizaje de habilidades conductuales) y la cognición (aprendizaje de habilidades cognitivas).Utilizando unos criterios, se han identificado cinco sistemas de memoria: El sistema de memoria de trabajo, el sistema de memoria procedimental, el sistema de representación perceptual, el sistema de memoria semántica, y el sistema de memoria episódica.
El concepto de sistemas de memoria es uno de los constructos fundamentales de la neurociencia cognitiva. La existencia de sistemas de memoria está bien asentada en la psicología cognitiva y la neurociencia, la idea tiene una larga historia pero sobre todo se convirtió en un tema objeto de intensa investigación a partir de los años ochenta y sobre todo de los años noventa del siglo XX, numerosas evidencias empíricas obtenidas a partir de estudios experimentales realizados con participantes sanos, pacientes amnésicos y en estudios realizados con animales de laboratorio apoyan la existencia de distintos sistemas de memoria.
El descubrimiento de las técnicas de imágenes cerebrales en los años setenta (la tomografía de emisión de positrones-PET, y después las imágenes por resonancia magnética- MRI) permitió a los investigadores hacer realidad lo que hasta entonces solo había sido el sueño de Pavlov. Estas técnicas de imágenes cerebrales permiten observar, cada vez con más detalle, el funcionamiento del cerebro y mapear los cambios locales de la circulación y el metabolismo cerebral asociados con los cambios en la actividad celular del cerebro. Estas técnicas han originado, además, el nacimiento de la neurociencia cognitiva.
GLOSARIO
• Amígdala. Pequeña estructura nerviosa del tamaño de una almendra, situada debajo de cada lóbulo temporal, directamente implicada en la emoción, las fobias y la ansiedad.
• BOLD. Siglas en inglés del término blood-oxigen-level dependence. Es el contraste de la imagen por resonancia magnética de la sangre desoxigenada descubierto por el Dr. Seiji Ogawa en 1990. Las neuronas carecen de reserva de energía en forma de glucosa y oxígeno. A través de la llamada respuesta hemodinámica, la sangre las deja glucosa para que puedan generar la energía que necesitan. Esto hace que haya un aumento de oxihemoglobina en las venas de la zona y un cambio de la razón local de oxihemoglobina a deoxihemoglobina, el marcador de BOLD. La señal de resonancia magnética de la sangre es ligeramente diferente dependiendo del nivel de oxigenación. Mayores intensidades de la señal BOLD surgen por el aumento en la concentración de hemoglobina oxigenada ya que la susceptibilidad magnética se parece más a la susceptibilidad magnética del tejido. Los cambios pueden ser positivos o negativos dependiendo de los cambios relativos en el flujo sanguíneo cerebral y en el consumo de oxígeno. La diferencia de la señal es muy pequeña, pero mediante muchas repeticiones (un gran número de ensayos) pueden utilizarse métodos estadísticos para ver qué áreas cerebrales muestran diferencias significativas como resultado de que se encuentran activas durante la realización de la tarea. Prácticamente toda la investigación actual en resonancia magnética funcional utiliza el método BOLD para determinar el lugar del cerebro donde se produce la actividad como resultado de la realización de la tarea que pone en marcha un determinado proceso psicológico.
• Cortezas parahipocámpicas y perirrinales. Son zonas cerebrales que rodean al hipocampo implicadas en la memoria.
• Demencia tipo Alzheimer. Enfermedad neurológica de tipo degenerativo que afecta a la memoria y a otras funciones cognitivas con un mantenimiento al menos durante las primeras etapas de las funciones sensoriales y motoras. Esta enfermedad fue descrita por primera vez el neurólogo alemán Alois Alzheimer.
• Diencéfalo. Es la parte del encéfalo situada entre el telencéfalo y el mesencéfalo o cerebro medio. Se trata de un abultamiento del compartimiento mediano más anterior del tubo neural embrionario.
• Función hemodinámica del cerebro. Flujo sanguíneo dentro del sistema vascular cerebral.
• Hipocampo. Estructura curva situada en el interior del cerebro, debajo del lóbulo temporal, de la que depende la memoria explícita. Se llama así porque tiene la forma de un caballito de mar. La lesión bilateral en esta zona cerebral produce amnesia.
• Hipnosis. Estado mental generado mediante un procedimiento conocido como inducción hipnótica que consiste en una serie de instrucciones y sugerencias que pueden ser generadas por el hipnotizador o ser autogeneradas. La hipnosis se utiliza para usos terapéuticos.
• Neurociencia cognitiva. Saber multidisciplinar integrado por una serie de ciencias como la química-física, la biofísica, la bioquímica, la anatomía, la neuroanatomía, la psiquiatría y la psicología. Su objeto es estudiar lo cognitivo a través de las ciencias biofísica y bioquímicas para lo cual se centran en la estructura y la función del Sistema Nervioso.
• Priming neural. Reducción de la actividad neural en varias regiones cerebrales como consecuencia de la repetición de estímulos. También se denomina supresión de repetición.
• Procesamiento guiado por los datos. También denominado procesamiento abajo-arriba porque está guiado por los aspectos físicos o sensoriales de los estímulos.
• Procesamiento guiado por el significado. También denominado procesamiento arriba-abajo o guiado por el significado.
• Región parahipocampal. Conjunto de áreas corticales que rodea al hipocampo, incluyendo la corteza perirrinal, la corteza entorrinal y la corteza parahipocampal.
• Resonancia magnética funcional (fMRI). Técnica hemodinámica que mide los cambios metabólicos producidos en una determinada parte del cerebro cuando se activa. Esta técnica de imágenes cerebrales utiliza ondas electromagnéticas de radio-frecuencia emitidas por el núcleo de los átomos de hidrógeno para construir imágenes detalladas del cerebro. Es una herramienta muy poderosa para estudiar los procesos cognitivos que ocurren en el cerebro humano.
• Sistemas de memoria. Postura teórica de la neuropsicología y la psicología cognitiva que mantiene que en el cerebro humano existen varios sistemas de memoria a largo plazo para explicar las disociaciones encontradas entre las distintas pruebas de memoria en personas sanas y en la amnesia.
• Sistema temporal medial diencefálico. Se denomina así una serie de áreas cerebrales situadas en la parte media del sistema temporal de las que depende la memoria episódica.
• Teorías del procesamiento de la información. Enfoque en el estudio de la psicología cognitiva surgido dentro de la tradición experimental norteamericana en la mitad del siglo pasado. Sus proponentes argumentan que igual que el ordenador digital, la mente humana es un sistema que procesa información mediante la aplicación de las reglas lógicas. Igual que ocurre con el ordenador, la mente humana tiene una capacidad limitada para procesar información.
• Voxel. Palabra inglesa correspondiente a pixel volumétrico que es la unidad cúbica que compone un objeto tridimensional. Es la unidad mínima procesable de una matriz tridimensional. Es equivalente al pixel en un objeto bidimensional (2D).
REFERENCIAS
- • Resumen Alejandra Mendieta Rada
- • Resumen Laura Jerónimo Olaya
- • Ballesteros Jiménez, S. (2017). Psicología de la memoria humana. Madrid: Universitas.
